Fig. 1
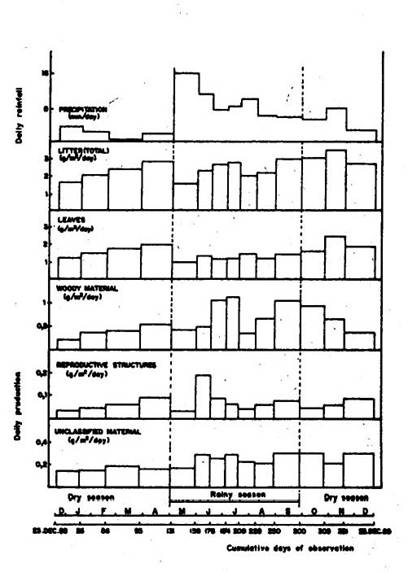
Fig. 2
The text that follows is a PREPRINT.
Please cite
as:
ISSN: 0564-3295
Copyright: International Society for Tropical Ecology (ISTE)
Carbon and nutrient flows in an Amazonian forest: fine litter production and composition at Apiaú, Roraima, Brazil
REINALDO IMBROZIO BARBOSA* AND PHILIP M. FEARNSIDE**
* Núcleo de Pesquisa de Roraima,
Instituto Nacional de Pesquisas da Amazônia (INPA), C.P. 96, 69301-970 Boa
Vista, Roraima, BRAZIL; ** Coordenadoria de Pesquisas em Ecologia, Instituto
Nacional de Pesquisas da Amazônia (INPA), C.P. 478, 69011-970 Manaus, Amazonas,
BRAZIL
** (Please address correspondence regarding this paper to the second author)
Running headline: FINE LITTER IN AN AMAZONIAN FOREST
17 July 1995
6 Dec. 1995
Carbon and nutrient flows in an Amazonian forest: fine litter production and composition at Apiau, Roraima, Brazil
R.I. Barbosa and P.M. Fearnside
Abstract: Production and nutrient composition of litter in an Amazonian tropical moist terra firme (upland) forest in Roraima, Brazil, was recorded over 12 months during 1988-89. Total production was 9.17 t ha-1 year-1, composed of 5.57 t ha-1 year-1 of leaves, 2.43 t ha-1 year-1 of woody material, 0.28 t ha-1 year-1 of reproductive structures, and 0.89 t ha-1 year-1 of unclassified material. Of the total litter, carbon represented 45.6%. Concentrations of nitrogen (1.51%), calcium (0.61%), and magnesium (0.15%) are similar to those of other tropical forest reports in Brazilian Amazonia. However, concentrations of phosphorus (0.06%) and potassium (0.44%) are among the highest levels recorded in the region. Production of potassium in the leaf fraction and in the total litter, and phosphorus in the leaves, are negatively correlated with precipitation. Mean daily production of leaves differs significantly between wet and dry seasons. Nitrogen and carbon produced in the leaf fraction also vary between wet and dry seasons, as well as calcium in the woody fraction and in unclassified material. A positive correlation exists separately between total litter production and leaf fractions, woody material and unclassified material.
Les flux de
carbone et d'éléments nutritifs dans une forêt Amazonienne: la production et
composition de la litiere fine a Apiau, Roraima, Brésil
R.I. BARBOSA et P.M. FEARNSIDE
Résumé: La production et composition des
éléments nutritifs de la litière dans une forêt tropicale Amazonienne de terre
firme à Roraima, Brésil, ont été mesurées pendant 12 mois (1988‑89). La production totale était de 9.17 t ha-1
an-1, composée de 5.57 t ha-1 an-1 de feuilles,
2.43 t ha-1 an-1 de bois, 0.28 t ha-1 an-1
de structures reproductrices, et 0.89 t ha‑1 an-1
de matière non classifiée. De la litière
totale, le carbone a représenté 45.6%.
Les concentrations d'azote (1.51%), de calcium (0.61%), et de magnésium
(0.15%) sont semblables à celles d'autres études en forêt tropicale de
l'Amazonie Bresilienne. Les
concentrations de phosphore (0.06%) et de potassium (0.44%) sont parmis les
plus‑hauts niveaux enregistrés dans la région. La production de potassium dans la fraction
des feuilles et dans la litière totale, et le phosphore des feuilles, montrent
une corrélation negative avec le niveau de précipitation. La moyenne quotidienne de production de
feuilles est significativement différente entre la saison sèche et humide. L'azote et le carbone, produits dans la
fraction de feuilles varient aussi entre la saison sèche et humide, bien comme
le calcium dans la fraction du bois et dans le matière non classifiée. Une corrélation positive existe separadamente
entre la production de la litière totale et les fractions de feuilles, bois et
matière non classifiée.
Flujos
de carbon y de nutrientes en una floresta amazonica: produccion y composicion de litera en Apiau,
Roraima, Brasil
R.I. BARBOSA y P.M. FEARNSIDE
Resumen: La producción y la composición mineral de la
litera en una floresta de tierra-firme en la región de Roraima, Brasil, fui
medida durante 12 meses en 1988-89. La
producción total era de 9.17 t ha-1 año-1, compuesta de
5.57 t ha-1 año-1 de hojas, 2.43 t ha-1 año-1 de material leñoso, 0.28 t ha-1
año-1 de estructuras reproductivas, y 0.89 t ha-1 año-1
de material no clasificado. Del total de
la litera, el carbón representó 45.6%.
Las concentraciónes de nitrógeno (1.51%), cálcio (0.61%), y magnésio
(0.15%) son semejantes a los valores de otras florestas tropicales en la
Amazonía brasileña. Mientras tanto, las
concentraciones de fósforo (0.06%) y de potássio (0.44%) estan entre los
niveles más altos registrados en la región.
La producción de potássio en la fracción foliar y en el total de la
litera, y el fósforo en las hojas, son negativamente correlacionados con la
precipitación. La producción média
diária de hojas difiere significtivamente entre la estación seca y lluviosa. Nitrógeno y carbón producidos en la fracción
de hojas también varían entre las estaciónes secas y lluviosas, así como cálcio
en la fracción leñosa y en el material no clasificado. Una correlación positiva existe separadamente
entre la producción total de la litera y las fracciones para hojas, material
leñoso y material no clasificado.
Fluxos de
carbono e de nutrientes numa floresta amazônica: produção e composição de liteira em Apiau,
Roraima, Brasil.
R.I.
BARBOSA e P.M. FEARNSIDE
Resumo: A produção e a composição mineral da liteira
em uma floresta de terra firme na região de Roraima, Brasil, foi medida durante
12 meses em 1988-89. A produção total
era de 9,17 t ha-1 ano-1, composta de 5,57 t ha-1
ano-1 de folhas, 2,43 t ha-1 ano-1 de material lenhoso, 0,28 t ha-1
ano-1 de estruturas reproductivos, e 0,89 t ha-1 ano-1
de material não classificado. Do total
da liteira, o carbono representou 45,6%.
As concentrações de nitrogênio (1,51%), cálcio (0,61%), e magnésio (0,15%)
são semelhantes aos valores de outras florestas tropicais na Amazônia
brasileira. No entanto, as concentrações
de fósforo (0,06%) e potassio (0,44%) são entre os níveis mais altos
registrados na região. A produção de
potássio na fração foliar e no total da liteira, e o fósforo nas folhas, são
negativamente correlacionados com a precipitação. A produção média diária de folhas difere
significtivamente entre a estação seca e chuvosa. Nitrogênio e carbono produzidos na fração
foliar também variam entre as estações secas e chuvosas, assim como cálcio na
fração lenhosa e no material não classificado.
Uma correlação positiva existe separadamente entre a produção total da
liteira e as frações para folhas, material lenhoso e material não classificado.
Keywords: Amazonia, Carbon cycling, Forest ecosystems, Litter production, Nutrient cycling, Tropical forest, Rainforest, Roraima
INTRODUCTION
Quantifying litter production and composition is an important area of ecological research in tropical forests (Proctor 1983). On the soil surface, the litter layer functions as an input-output system, receiving large quantities of plant detritus (Ewel 1976). This process is of fundamental importance for the understanding of ecosystem functioning because litter fall is the largest pathway for nutrient and organic matter flux to the soil (Meentemeyer et al. 1982).
Human occupation in Amazonia is rapidly changing the land-use patterns in the region. Felling and burning of forest for agriculture or cattle pasture changes the nutrient cycling pattern in natural ecosystems, changing the equilibrium between production of biomass, accumulation of organic matter and decomposition and absorption of minerals. This frequently results in nutrients becoming a limiting factor for crop yields when tropical forests are converted to agricultural use (Jordan 1985).
Reviews done by Proctor (1984) and Vitousek (1984) indicate that the number of studies on litter in Brazilian Amazonia is small compared with other tropical areas. In spite of its importance, most of the work in this region only began in the late 1970s, with the studies of Klinge (1977a,b), Adis et al. (1979), Franken (1979), Franken et al. (1979), Silva & Lobo (1982), Silva (1984), Luizão & Schubart (1987), Dantas & Phillipson (1989) and Luizão (1989). These studies report on various types of vegetation, and are restricted to the areas of Manaus and eastern Amazonia (especially the Belém area). In Roraima, the northernmost part of the Brazilian Amazon, the only study is that of Scott et al. (1992), carried out in 1987-1988 at the Maracá Ecological Station.
The present study is intended to quantify the production and mineral composition of litter in an area of terra firme tropical forest in Roraima. Relationships will be worked out between litter biomass production, nutrient flux and rainfall, and deposition of carbon in the soil. This information is essential to any effort to model nutrient flows in these forests, with or without the effects of human perturbation.
MATERIALS AND METHODS
Study area
The study area is located in Colônia do Apiaú (2o34' N, 61o18' W), a government colonization project in the município of Mucajaí, state of Roraima, Brazil. The area is a part of the "Projeto de Colonização Rio Apiaú", covering 23,608 ha in which settlement began in November 1981 (GTFR 1980a,b,nd). It is estimated that in 1993 the area of the colonization project has increased to about 60,000 ha, which represents a growth of approximately 250% in 12 years.
The area is drained by the Apiaú and Mucajaí rivers. The major topographic features are two ranges of hills: the Serra de Mucajaí to the south, and the Serra do Ipiranga to the north. The altitude varies between 100 and 180 m (excepting the hills). The climate is classified as "Ami" in the Köppen system, a rainy tropical climate with a marked dry season (January to March) (Santos et al. 1982). The mean annual rainfall is approximately 2000 mm, based on rainfall at locations near the colonization project (Lameira & Coimbra 1988, and the present study), and the relative humidity of the air has an annual mean of 87% (GTFR 1980a).
The soils in the region are generally acidic and poor in nutrients. Under dense forest at this site, the pH varied from 3.66 to 4.88, and the concentration of available phosphorus was 2.5 ppm in the top 10 cm of the soil (Barbosa 1991). The predominant soil types in this location are red-yellow podzolic (Ultisol), red-yellow latosol (Oxisol), and the lithosols that occupy the mountainous parts of the region (Brazil, Projeto RADAMBRASIL 1975).
The natural vegetation is predominantly terra firme tropical forest, which according to the side-looking airborne radar (SLAR) maps of the RADAMBRASIL Project (Brazil, RADAMBRASIL 1975) fall into the following categories: (1) area of ecological tension of contact between forest and seasonal forest, and (2) dense tropical forest of low chains of mountains. The present study area is located in a 2.5 km2 block of the second type, where the predominant soil is red-yellow podzolic (Ultisol).
According to a commercial timber survey by the Roraima government (GTFR 1980c), the main species are casca grossa (Qualea paraensis), abiurana (Pouteria spp.), angelim (Dinizia excelsa), fava-benguê (Parkia spp.), cupiúba (Goupia glabra), rabo-de-arraia (Qualea cf. brevipedicellata), maçaranduba (Manilkara spp.), breu (Protium spp.), jará (Sarcaulis brasiliensis), louro branco (Ocotea opifera), ripeiro (Eschweileira amara) and piquiá (Caryocar spp.). In addition to these, the presence of angelim-ferro (Hymenolobium spp.) is noteworthy, this species appearing as an emergent in monospecific stands in some parts of the forest. Jatobá (Hymenaea spp.) is also characteristic. Two palms, inajá (Maximiliana maripa) and açaí (Euterpe spp.), although not listed in the government survey, can be seen in substantial numbers in the settlement area. According to the government survey, the density of trees with a diameter at breast height (DBH) of 40 cm or more in this forest type is 18.3 trees ha-1, with a trunk volume below the first branch of 45.9 m3 ha-1.
Collection and preparation of samples
Six 1 X 1 m wooden boxes were installed lined with plastic sheeting with a hole in the center for litter collection. The boxes were suspended at a height of 1.2 m above ground level, on top of empty 200-liter oil drums which served as pluviometers for another experiment set up at the same site. A plastic bucket was suspended under each box catching material that washed through the hole in the plastic sheet.
Two blocks of three boxes were set up, the groups being 100 m apart within the forest. The boxes within each group were set up in a triangular pattern, with each box 15 m from the others.
Litter collection was carried out from 17 August 1988 to 23 December 1989, at intervals of approximately 6-10 days. The 23 December 1988-23 December 1989 period values are presented here, as the detritus was not collected in the plastic buckets in the period prior to this.
All materials collected were dried in an electric oven at 75oC for 72 hours, followed by manual sorting of the material into the following pre-established fractions: leaves, reproductive structures (fruits, flowers, seeds), woody material (bark and wood--branches and twigs with diameter <2.5 cm), and unclassified material (material not included in the other fractions: insects, fecal matter, etc.). After drying and sorting, each fraction was weighed on an electronic balance. Dry weights were determined for each fraction in each collection.
Each group of three successive collections was lumped to form a sample, yielding a total of 14 samples representing periods which varied from 15 to 35 days. The material was milled and analyzed by R.I.B. in the Laboratory for Analysis of Soil and Plant Tissues of the Brazilian Enterprise for Agricultural and Cattle Ranching Research--Territorial Level Experimental Unit (EMBRAPA-UEPAT) in Boa Vista.
Chemical analyses
An analysis of nutrient concentrations was done for the following elements in each fraction of the sample: N, P, K, and Ca & Mg. Before formal processing for routine analysis, ground material was first placed in an oven for 24 hours at 50oC in order to standardize the humidity content among the different fractions. About 0.2 g was then weighed out from each sample for the process of digestion by H2O2 and H2SO4, following the method described by Tedesco et al. (1985). The extract so obtained was used for determining the elements using the following methods: N by acid titration with H2SO4 (0.05 N) in a Kjeldahl distillation apparatus; P by colorimetry in a spectrophotometer (Micronal, model B380); K by flame photometry (Micronal, model B262); Ca & Mg by atomic absorption spectrophotometry (Perkin/Elmer, model 2380).
For determination of total carbon, a subsample of 0.1 g of ground plant tissue was weighed out and analyzed by the Tedesco et al. (1985) method. This method is designed for analysis of plant samples, and is a modification of the Walkley-Black method (Allison 1965) of soil carbon determination.
Statistical treatment of the data
Pearson's correlation coefficient was used (Zar 1974) to establish the relationship between precipitation and the production of nutrients and carbon. Analysis of variance (ANOVA) was employed to test the difference between the means encountered in the dry and rainy seasons, both for production of litter biomass and for nutrients and carbon.
RESULTS
Seasonality and production of litter fractions
The total litter production rate was 9.17 t ha-1 year-1 (Table 1). There was no significant difference between average daily production during the dry season (2.6 g m-2 day-1) and the rainy season (2.4 g m-2 day-1). In the driest part of the year (January to March), the leaf fraction contributed 72.5% to total litter production. The production of woody material increased during the rainy season (May to mid-September), having an average value of 32% of total litter production. Reproductive structures maintained an even production during the year, with an exception of a peak in June, soon after a rainy spell of 15.9 mm day-1. Unclassified material showed the greatest proportion during the rainy season, with an average of 10.9%.
(Table 1 here)
The greatest production of total litter occurred between September and November, i.e. in the transition phase between the wet and dry seasons. The peak production at this time was 3.5 g m-2 day-1, coinciding with the greatest production of leaves, 2.5 g m-2 day-1. There was a secondary peak of litter production at the end of the dry season between March and April (Figure 1).
(Figure 1 here)
The production rate of the leaf fraction showed significantly different means (ANOVA, P< 0.01) in the rainy season (1.3 g m-2 day-1) and in the dry season (1.8 g m-2 day-1). Woody material did not show significant differences in production rate between the dry season (0.5 g m-2 day-1) and the wet season (0.8 g m-2 day-1), even though the numerical difference appears marked.
The unclassified material category also had the greatest production rate in the rainy season (0.3 g m-2 day-1). However, there was no significant difference between the two seasons. The behavior of reproductive structures was similar, also showing the greatest production during the wet season (0.09 g m-2 day-1), but not significantly different from the dry season rate (0.07 g m-2 day-1).
An analysis of variance among the collection boxes revealed a coefficient of variation of 34.8%, when analyzed in terms of total production. Leaves presented the least coefficient with 28.0%, followed by unclassified material (50.7%), reproductive structures (67.2%) and woody material (69.8%).
Carbon and nutrient fluxes
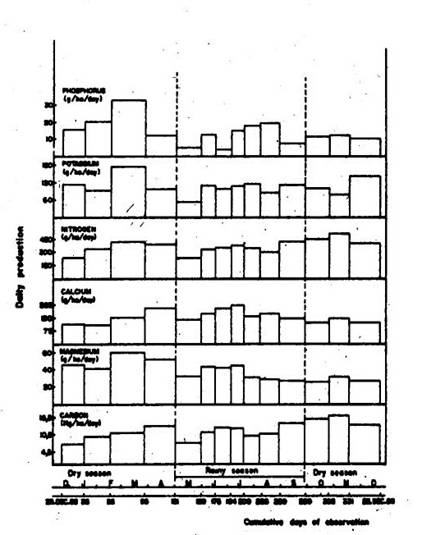
The nutrients with the greatest proportion were nitrogen (138 kg ha-1 year-1) and calcium (56 kg ha-1 year-1), followed by potassium (40 kg ha-1 year-1), magnesium (14 kg ha-1 year-1) and phosphorus (6 kg ha-1 year-1). The carbon production represented 45.6% of total litter production (Table 2).
(Table 2 here)
Phosphorus, potassium and magnesium had their maximum productivity at the height of the dry season (33, 173 and 60 g ha-1 day-1, respectively) (Figure 2). The greatest production of nitrogen (523 g ha-1 day-1) was at the same time as the peak in litter production (October-November). Calcium showed its maximum production (225 g ha-1 day-1) in the rainy season, coinciding with the major fall of woody biomass. At the end of the rainy season, carbon had its greatest production (17 kg ha-1 day-1), the same as nitrogen. The production of these two elements is significantly associated in all fractions analyzed, especially in leaves (r=0.942, P< 0.001) and woody material (r= 0.980, P< 0.001). In total litter the behavior was the same (r= 0.929, P< 0.001).
(Figure 2 here)
The mean carbon production in the leaf fraction showed a significant difference between the two seasons (ANOVA, P< 0.01). The same occurred with nitrogen (ANOVA, P< 0.05). Calcium also showed a significant difference between averages obtained in the wet and dry seasons, for woody material (ANOVA, P< 0.05), and unclassified material (ANOVA, P< 0.01).
The C/N ratio averaged 32 over the period studied. The fraction that had the greatest numerical value was woody material (42), followed by leaves (32), reproductive structures (28), and unclassified material (22). The highest ratio found during the entire collection period was in July (in the rainy season) in the woody material fraction (51). This ratio coincided with the greatest daily production in this fraction and in Ca. The lowest was in January-February (dry season) for unclassified material (16).
DISCUSSION
Litter production
Litter production recorded in this study was among the highest known in Amazonian Brazil when compared to previous studies in the region. The results are equivalent to those obtained by Klinge (1977b) in Belém, and Scott et al. (1992) in Roraima. The results obtained are also similar to those for terra firme forests elsewhere in Amazonia and neighboring parts of South America (Table 3).
(Table 3 here)
Litter production was greatest in the dry season, due to the greater number of days considered for this season in the year. Mean daily production in this season was also slightly higher in comparison with the rainy season. This tendency was similar to the findings of Klinge & Rodrigues (1968a), Dantas & Phillipson (1989), Luizão (1989) and Scott et al. (1992). Even with correction of the daily means, this seasonal model of production showed itself to be unpredictable in our study, due to the great difference in the days counted for rainy and dry seasons.
The greatest daily productions of total litter occurred between the end of the rainy season and the beginning of the dry season. This period represents the beginning of a phase of high temperatures associated with torrential rains and strong winds. These storms are usually interspersed with longer rainless periods (8-23 days) when the peak of the dry season draws closer. This period is also characterized by higher insolation, with a mean of 205 hours month-1 between September and March, a value 1.37 times higher than the mean for the remainder of the year (Brazil, Instituto Nacional de Meteorologia 1980). The combination of these climatic factors could cause a greater litterfall rate during this period due to (1) physiological stimulation, with shedding of senile material to prepare for future investment in vegetative growth, or (2) mechanical breaking of plant parts.
Although a correspondence may exist, it would be tendentious to explain litter production by instantaneous climatic factors. Bazilevich et al. (1971) suggest that phases with characteristics such as longer photoperiod and higher relative humidity can cause more rapid growth. However, in order for this to occur, it is necessary to have other factors or physiological activities that stimulate the fall of senile material from the plant and regulate the activation of growth functions. Edaphic factors and decreasing rainfall could activate this mechanism. In part, this could explain the high rates of litter production at the beginning of the dry season in this region. On the other hand, some authors have observed increases in litterfall associated with winds (John 1973, in Ghana, and Alvarez-Sánchez & Seda 1993, in Mexico) and storms (Luizão 1989, in Manaus). The impact of rainfall intensity could have a greater ecological significance than the annual and monthly means for precipitation (Brinkmann 1985), and could be a decisive factor in the fall of detritus.
The lowest production of litter was found at the beginning of the driest part of the year (January), associated with high rates of insolation. In tropical areas, the maximum use of solar radiation tends to be of great importance for primary productivity (Bray & Gorham 1964), resulting in the plant making a greater investment in vegetative growth. This association between the availability of light and litter production was also emphasized by Jordan (1971). This could indicate that low levels of precipitation do not necessarily cause an associated production of detritus. This parameter also should be accompanied by low indices of soil humidity. At our study site, this fact could have stimulated the secondary peak of production, probably associated with water stress at the end of the dry season (March-April).
The leaf fraction was responsible for a high percentage of the total litter production, as has also been found in other studies carried out in Amazonia. This fraction, when disaggregated from total litter and analyzed separately, showed a strong seasonal influence. This corroborates the hypothesis of Wright & Cornejo (1990) that greater production of leaves is associated with the dry season in tropical forests at low latitudes (<13o) and altitudes (<600 m).
Folster & de las Salas (1976) found in Colombia that production of woody material coincided with the maximum peak of rainfall. These authors sampled woody material to a maximum diameter of 3.5 cm. The present study found similar results, but used a maximum diameter of 2.5 cm.
The greatest production of unclassified material occurred in the rainy season. This suggests a greater biological activity on the detritus deposited in the boxes. Intense washing of the tree crowns, in addition to an increase in fecal matter and dead insects at this time of year, would also contribute to this result.
The reproductive structures showed an irregular cycle, and influence from seasonal variations was not evident. The same phenomenon was observed by Cuevas & Medina (1986) in San Carlos de Río Negro in Venezuela, and is probably due to the differences in species found in different tropical forests and their different phenological rhythms (Fassbender & Grimm 1981).
The high coefficients of variation found for woody material, reproductive structures and unclassified material indicate the great natural variability in the production of these components. It could also be a result of the small number of collection boxes used in the study. However, in the study conducted by Scott et al. (1992) on Maracá Island, approximately 95 km from the study area, a production of 9.28 t ha-1 year-1 was found, similar to the results of the present study, using a greater number of smaller collectors. A greater number of collectors increases the representativeness of the microsites sampled, but smaller collectors preclude a representative sample of the larger pieces of detritus. The representativeness of the study area, the number of collectors, and the area of each collector are important parameters that still need better definition in litter studies. One way in which litter sampling methods often differ is the time between collections. Our study, with a mean interval of 8 days and a range of 4-17 days (with shorter intervals in rainy periods), has less bias from decomposition of litter in the traps prior to collection than do many studies with intervals of 30 days or more. Our samplers also lose less fine material than do samplers constructed of meshes, often with openings of 2 mm or larger. Studies often differ in the definition of fine litter (defined in the present study by a maximum diameter of 2.5 cm).
Carbon and nutrient fluxes
The concentration of phosphorus (0.06%) for this area is higher than most measurements in terra firme forests of Brazilian Amazonia. Of the seven existing studies with at least one year of collection in Brazilian Amazonia (Table 3), only the other study conducted in Roraima (Scott et al. 1992) found a higher value (0.072%). In a half-year study near Manaus, Stark (1971) found a value of 0.061%, but the seasonal nature of high-phosphorus components such as reproductive structures makes this value not comparable. In low-altitude terra firme forests elsewhere, Poels (1987), citing Ohler (1980), also reports a high concentration (0.065%) in Suriname but the material was collected from the soil surface rather than in traps. In Panama, Golley et al. (1975) found a concentration of 0.074%. In other forest types (other than mature low-altitude terra firme evergreen tropical forests), values equal to or greater than ours have been found: Fassbender & Grimm (1981) found 0.057% in montane forest in Venezuela, Veneklaas (1991) found 0.087% in montane forest in Colombia, Nadkarni & Matelson (1992) found 0.096% in montane cloud forest in Costa Rica, and Ewel (1976) found 0.064% in old secondary forest in Guatemala.
The concentration and production of nitrogen found in the present study corroborate the hypothesis that this element is not a limiting factor in tropical forests (Vitousek 1982). The relative abundance of legumes found in these forests, potentially responsible for symbiotic fixation of atmospheric nitrogen, supplies high levels of this element at these sites (Jenny 1950; Vitousek 1984). Nye (1961) reported that, in general, tropical forests have higher concentrations of nitrogen than comparable temperate forests, due to the higher concentration of this element found in the leaf fraction. However, our study indicates a greater concentration of this nutrient in the reproductive structures and unclassified material fractions, although leaves were responsible for 64% of production of this bioelement.
The highest concentration of calcium was found in the woody material fraction, as has also been found in various other studies. In addition to this, seasonality of production of this nutrient as a function of production of this litter fraction and of unclassified material was marked.
High levels of phosphorus and potassium in reproductive structures suggests that this litter component concentrates these elements. The total production of potassium was also one of the highest found so far in Amazonia. The short period between collections (6-10 days) could have influenced this high concentration in comparison to other studies (see Table 3). A short time exposed in the collection boxes would lead to low losses of this element, which is easily leached out. However, this value can represent a higher level of potassium in the vegetation, due to the differences in the pedological formation between, for example, central-west Amazonia and the Roraima area. It is probable that the concentration of nutrients in the different plant tissues reflects the fertility differences in the soil (Vitousek & Sanford 1986).
Klinge & Rodrigues (1968b), in a study carried out near Manaus, found high concentrations of phosphorus and potassium in the litter in the rainy months, but the greatest production of these elements was in the dry season, when the rate of litterfall is greatest. This pattern was found in the present study only for the leaf component of the litter, as the other components had production and/or concentrations in the opposite direction from that found by Klinge & Rodrigues (1968b).
The production of carbon had the same pattern of concentration as in the studies carried out near Manaus. The significant difference found between the means for production and the two periods of the year merely reflects the greater total volume of litter produced and the difference also observed for the leaf fraction. Both showed similar fluctuations in production during the year.
The average C/N ratio of litter fractions found in this study is of the same order as that observed in other studies in tropical forests. The higher values encountered for woody material and leaves indicate that, beginning with the time of deposition on the ground, microorganisms will have the most difficulty in degrading these fractions. Average C/N ratios above 20 would indicate slow decomposition with immobilization of N. The inverse applies to unclassified material and reproductive structures. These fractions have C/N ratios that provoke rapid degradation and stimulate decay processes.
In our study no large accumulation of leaves was observed in the litter layer on the soil, indicating that the higher C/N ratio for this material was not an impediment to rapid decomposition. However, the presence of woody material was readily apparent anywhere on the forest floor. Different compositions of the litter layer on the soil surface influence the community of microbial decomposers present throughout the year, with the predominant material defining the presence of different groups (Puig & Delobelle 1988).
CONCLUSIONS
The results of this study confirmed many of the qualitative and quantitative characteristics found in other tropical forest sites. They also show important differences, such as (1) one of the highest production rates of total litter known in Brazilian Amazonia, (2) a peak of litter production at the end of the rainy season, suggesting that shedding of senile material is being stimulated by the beginning of the period of greater insolation and of higher soil humidity, (3) a secondary peak of litter production associated with water stress at the end of the dry season, and (4) high concentrations of phosphorus and potassium, which are not common in other parts of the Brazilian Amazon. These differences indicate the need for studies with a long time series, and for increasing the number and diversity of sites to be studied in the very heterogeneous forests of Amazonia.
ACKNOWLEDGMENTS
The government of Roraima funded part of this study through Convênio No. 072/85 (Governo do Território Federal de Roraima/ Instituto Nacional de Pesquisas da Amazônia-GTFR/INPA). We thank the Empresa Brasileira de Pesquisa Agropecuária-Unidade Experimental no Ambito Territorial--EMBRAPA-UEPAT), Boa Vista, for permitting us to analyze the samples in their laboratories. We also thank Manoel Dorneles de Souza and Regina Martoni Mansur Corrêa da Costa who helped, respectively, with the laboratory and field activities. C. Gonçalo recorded the daily rainfall in the Colônia de Apiaú between 1988 and 1989. Flávio Luizão and S.V. Wilson made valuable comments on the manuscript.
REFERENCES
Adis, J., K. Furch & U. Irmler. 1979. Litter production of a Central-Amazonian black water inundation forest. Tropical Ecology 20(2): 236-245.
Allison, L.E. 1965. Organic Carbon. pp. 1367-1379 In: C.A. Black (ed.) Methods of Soil Analysis. Agronomic Series No. 9. Agronomic Society of America, Madison, Wisconsin.
Alvarez-Sánchez, J. & S.G. Sada. 1993. Litterfall dynamics in a Mexican
lowland tropical rain forest. Tropical Ecology 34(2): 127-142.
Barbosa, R.I. 1991. Erosão do solo na Colônia do Apiaú, Roraima,
Brasil: Dados preliminares. Boletim do Museu Integrado de Roraima 1(2):
22-40.
Bazilevich, N.I., A.V. Drozdov & L.E.
Rodin. 1971. World forest
productivity, its basic regularities and relationships with climatic factors.
pp. 345-353 In: Productivity of Forest Ecosystems. UNESCO, Paris.
Brazil, Instituto Nacional de Meteorologia
(INM). 1980. Boletim climatológico do
município de Boa Vista--1923 a 1979. Ministério da Agricultura, INM,
DFA/Roraima/1o DIEME, Boa Vista, Roraima, Brazil.
Brazil, Projeto RADAMBRASIL. 1975. Levantamento de Recursos Naturais, Volume
8. Ministério das Minas e Energia, Departamento Nacional de Pesquisa
Mineral (DNPM), Rio de Janeiro, Brazil.
Bray, J.R. & E. Gorham. 1964. Litter production in forest of the world. Advances in Ecological Research 2: 101-157.
Brinkmann, W.L.F. 1985. Studies on hydrobiogeochemistry of a tropical lowland forest system. GeoJournal 11(1): 89-101.
Cuevas, E. & E. Medina. 1986. Nutrient dynamics within Amazonian forest ecosystems. I. Nutrient flux in fine litter fall and efficiency of nutrient utilization. Oecologia 68: 466-472.
Dantas, M. & J. Phillipson. 1989. Litterfall and litter nutrient content in primary and secondary Amazonian 'terra-firme' rain forest. Journal of Tropical Ecology 5: 27-36.
Ewel, J.J. 1976. Litter fall and leaf decomposition in a tropical
forest succession in eastern Guatemala. Journal of Ecology 64: 293-308.
Fassbender, H.W. & U. Grimm. 1981. Ciclos biogeoquímicos en un ecosistema
forestal de los Andes occidentales de Venezuela. II. Produción y descomposición
de los resíduos vegetales. Turrialba 31(1): 39-47.
Folster, H. & G. de las Salas. 1976. Litter fall and mineralization in three
tropical evergreen forest stands, Colombia. Acta Cientifica Venezolana 27: 196-202.
Folster, H., G. de las Salas & P. Khana. 1976. A tropical evergreen forest site with perched water table, Magdalena Valley, Colombia. Biomass and bioelement inventory of primary and secondary vegetation. Ecologia Plantarum 11(4): 297-320.
Franken, M. 1979. Major nutrient and energy contents of the litterfall of riverine forest of Central Amazonia. Tropical Ecology 20(2): 211-224.
Franken, M., U. Irmler & H. Klinge. 1979. Litterfall in inundation, riverine and terra-firme forests of Central Amazonia. Tropical Ecology 20(2): 225-235.
Golley, F.B., J.T. McGinnis, R.G. Clements, G.I. Child & M.J. Duever. 1975. Mineral Cycling in a Tropical Forest Ecosystem. University of Georgia Press, Athens, Georgia.
GTFR - Governo do Território Federal de
Roraima. 1980a. Projeto de colonização
Rio Apiaú - Carta Consulta ao Banco Nacional de Desenvolvimento Econômico
(BNDE). Companhia de Desenvolvimento de Roraima/Secretaria de Planejamento
(CODESAIMA/SEPLAN-RR), Boa Vista, Roraima, Brazil.
GTFR - Governo do Território Federal de
Roraima. 1980b. Detalhamento de alguns
itens do projeto de colonização Rio Apiaú - Solicitação do Banco Nacional de
Desenvolvimento (BNDE). Secretaria de Planejamento (SEPLAN-RR), Boa Vista,
Roraima, Brazil.
GTFR - Governo do Território Federal de
Roraima. 1980c. Inventário florestal
exploratório do projeto Apiaú. Universidade Federal Rural do Rio de Janeiro
(UFRRJ), Instituto de Florestas, Boa Vista, Roraima, Brazil.
GTFR - Governo do Território Federal de
Roraima. nd. Relatório de avaliação
sobre o projeto Apiaú. Universidade Federal Rural do Rio de Janeiro (UFRRJ),
Instituto de Florestas, Boa Vista, Roraima, Brazil.
Jenny, H. 1950. Causes of the nitrogen and organic matter content of certain tropical forest soils. Soil Science 69: 63-69.
John, D.M. 1973. Accumulation and decay of litter and net production of forest in tropical West Africa. Oikos 24: 430-435.
Jordan, C.F. 1971. A world pattern in plant energetics. American Scientist 59: 426-433.
Jordan, C.F. 1985. Nutrient Cycling in Tropical Forest Ecosystems. Wiley, Chichester, U.K.
Klinge, H. 1977a. Preliminary data on nutrient release from decomposing leaf litter in a neotropical rain forest. Amazoniana 6(2): 193-202.
Klinge, H. 1977b. Fine litter production and nutrient return to the soil in three natural forest stands of eastern Amazonia. Geo-Eco-Trop 1(2): 159-167.
Klinge, H. & W.A. Rodrigues. 1968a. Litter production in an area of Amazonian terra-firme forest. I. Litterfall, organic carbon and total nitrogen contents of litter. Amazoniana 1(4): 287-302.
Klinge, H. & W.A.
Rodrigues. 1968b. Litter production in
an area of Amazonian terra-firme forest. II. Mineral nutrient content of the litter. Amazoniana 1(4):
303-310.
Lameira, O.A. & J.M. Coimbra. 1988. Levantamento e distribuição da precipitação
em Roraima. Empresa Brasileira de Pesquisa Agropecuária-Unidade Experimental de
Âmbito Territorial (EMBRAPA-UEPAT)/Boa Vista-Roraima. Pesquisa em Andamento
No. 12, Boa Vista, Roraima, Brazil.
Luizão, F.J. 1989. Litter production and mineral element input to the forest floor in a Central Amazonian forest. GeoJournal 19(4): 407-417.
Luizão, F.J. & H.O.R. Schubart. 1987. Litter production and decomposition in a terra-firme forest of Central Amazonia. Experientia 43: 259-265.
Meentemeyer, V., E.O. Box & R. Thompson. 1982. World patterns and amounts of terrestrial plant litter production. BioScience 32(2): 125-128.
Nadkarni, N.M. & T.J. Matelson. 1992. Biomass and nutrient dynamics of fine litter of terrestrially rooted material in a neotropical montane forest, Costa Rica. Biotropica 24(2a): 113-120.
Nye, P.H. 1961. Organic matter and nutrient cycles under moist tropical forest. Plant and Soil 13(4): 333-346.
Ohler, F.M.J. 1980. Phytomass and mineral content in untouched forest (CELOS Report 132). CELOS, Paramaribo, Suriname.
Poels, R.L.H. 1987. Soils, Water and Nutrients in a Forest Ecosystem in Suriname. Agricultural University, Wageningen, The Netherlands.
Proctor, J. 1983. Tropical forest litterfall. I. Problems of data comparison. pp. 267-273 In: S.L. Sutton, T.C. Whitmore and A.C. Chadwick (eds.) Tropical Rain Forest: Ecology and Management. Blackwell Scientific Publications, Oxford, U.K.
Proctor, J. 1984. Tropical forest litterfall. II. The data set. pp. 83-113 In: A.C. Chadwick & S.L. Sutton (eds.) Tropical Rain Forest: The Leeds Symposium. Leeds Philosophical and Literary Society, Leeds, U.K.
Puig, H. 1979.
Production de litière en forêt Guyanaise: resultates preliminaires. Bulletin
de la Societé d'Histoire Naturelle de Toulouse 115(3/4): 338-346.
Puig, H. & J.P. Delobelle. 1988. Production de litière, nécromasse, apports
minéraux ao sol para la litière en forêt Guyanaise. Revue d'Ecologie 43: 3-22.
Santos, R.D., J.N.R.F. Gama & A.F. Soares.
1982. Levantamento de reconhecimento
de baixa intensidade dos solos e avaliação da aptidão agrícola das terras do
projeto de colonização Apiaú, Território Federal de Roraima. Boletim de
Pesquisa No. 14. Empresa Brasileira de Pesquisa Agropecuária/Serviço Nacional
de Levantamento e Conservação de Solos (EMBRAPA/SNLCS), Rio de Janeiro, Brazil.
Scott, D.A., J. Proctor & J. Thompson. 1992. Ecological studies on lowland evergreen rain forest on Maracá Island, Roraima, Brazil. II. Litterfall and nutrient cycling. Journal of Ecology 80: 705-717.
Silva, M.F.F. 1984. Produção anual de serrapilheira e seu conteúdo mineralógico na mata
tropical de terra firme, Tucuruí-PA. Boletim
do Museu Paraense Emílio Goeldi, Série Botânica 1(1/2): 111-158.
Silva, M.F.F. & M.G.A. Lobo. 1982. Nota sobre deposição de matéria orgânica em florestas de terra firme,
várzea e igapó. Boletim do Museu Paraense Emílio Goeldi, Série Botânica 56:
1-13.
Stark, N. 1971. Nutrient cycling. II. Nutrient distribution in Amazonian vegetation. Tropical Ecology 12(2): 177-201.
Tedesco, J.M., S.J.
Volkweiss & H. Bohnen. 1985. Análises de Solo, Plantas e outros
Materiais. Departamento de Solos, Faculdade de Agronomia, Universidade
Federal do Rio Grande do Sul (DS/FA/UFRS), Boletim Técnico No. 5, Porto Alegre,
Rio Grande do Sul, Brazil.
Veneklaas, E.J. 1991. Litterfall and nutrient fluxes in two montane tropical rain forests, Colombia. Journal of Tropical Ecology 7: 319-336.
Vitousek, P. 1982. Nutrient cycling and nutrient use efficiency. American Naturalist 119(4): 553-572.
Vitousek, P. 1984. Litterfall, nutrient cycling and nutrient limitation in tropical forests. Ecology 65(1): 285-298.
Vitousek, P. & R.L. Sanford. 1986. Nutrient cycling in moist tropical forest. Annual Review of Ecology and Systematics 17: 137-167.
Wright, S.J. & F.H. Cornejo. 1990. Seasonal drought and leaf fall in a tropical forest. Ecology 71(3): 1165-1175.
Zar, J.H. 1974. Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, New Jersey.
FIGURE LEGENDS
Fig. 1. Daily production of litter fractions.
Fig. 2. Daily production of carbon and nutrients in the litter.
|
TABLE 1 ‑ Production
and Fractional Composition of Total Litter in an Amazon Forest in Roraima,
Brazil |
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||
|
Interval (dd/mm/yy) |
Days elapsed |
Leaves |
Wood |
Reproductive structures |
Unclassified material |
Total |
||||||
|
|
|
(g m-2 day-1) |
(%)* |
(g m-2 day-1) |
(%)* |
(g m-2 day-1) |
(%)* |
(g m-2 day-1) |
(%)* |
(g m-2 day-1) |
||
|
23/12/88‑17/01/89 |
26 |
1.20 |
74.4 |
0.22 |
13.7 |
0.04 |
2.4 |
0.15 |
9.5 |
1.62 |
||
|
18/01/89‑16/02/89 |
30 |
1.49 |
71.0 |
0.39 |
18.8 |
0.05 |
2.3 |
0.17 |
7.9 |
2.09 |
||
|
17/02/89‑27/03/89 |
39 |
1.74 |
72.0 |
0.43 |
17.8 |
0.05 |
2.2 |
0.20 |
8.1 |
2.42 |
||
|
28/03/89‑02/05/89 |
36 |
2.01 |
70.2 |
0.57 |
20.0 |
0.11 |
3.7 |
0.17 |
6.1 |
2.85 |
||
|
03/05/89‑30/05/89 |
28 |
0.98 |
59.4 |
0.46 |
27.5 |
0.04 |
2.4 |
0.18 |
10.6 |
1.66 |
||
|
31/05/89‑16/06/89 |
17 |
1.35 |
57.0 |
0.52 |
21.8 |
0.21 |
8.8 |
0.29 |
12.4 |
2.36 |
||
|
17/06/89‑04/07/89 |
18 |
1.17 |
43.6 |
1.15 |
42.6 |
0.10 |
3.6 |
0.27 |
10.2 |
2.69 |
||
|
05/07/89‑19/07/89 |
15 |
1.18 |
42.9 |
1.20 |
43.8 |
0.07 |
2.4 |
0.30 |
10.9 |
2.74 |
||
|
20/07/89‑07/08/89 |
19 |
1.46 |
68.2 |
0.39 |
18.0 |
0.05 |
2.4 |
0.24 |
11.4 |
2.13 |
||
|
08/08/89‑29/08/89 |
22 |
1.21 |
54.4 |
0.72 |
32.2 |
0.07 |
3.1 |
0.23 |
10.3 |
2.23 |
||
|
30/08/89‑28/09/89 |
30 |
1.43 |
47.7 |
1.17 |
38.8 |
0.09 |
2.9 |
0.32 |
10.7 |
3.01 |
||
|
29/09/89‑26/10/89 |
28 |
1.67 |
54.9 |
0.99 |
32.6 |
0.06 |
1.9 |
0.32 |
10.6 |
3.04 |
||
|
27/10/89‑18/11/89 |
23 |
2.50 |
71.4 |
0.71 |
20.1 |
0.07 |
1.9 |
0.23 |
6.6 |
3.51 |
||
|
19/11/89‑23/12/89 |
35 |
1.92 |
70.2 |
0.39 |
14.4 |
0.10 |
3.5 |
0.32 |
11.9 |
2.73 |
||
|
Production (g m-2
day-1) |
‑ |
1.52 |
|
0.66 |
|
0.08 |
|
0.23 |
|
2.51 |
||
|
Production (t ha-1
year-1) |
‑ |
5.57+1.61 |
2.43+1.59 |
0.28+0.18 |
0.89+0.44 |
9.17+3.19 |
||||||
|
Fracional composition(%) |
60.7 |
|
26.5 |
|
3.1 |
|
9.7 |
|
||||
|
* Percent
of each component in the total litter production during the period. |
|
|
|
|||||||||
|
TABLE 2. Total flux and concentration of elements in
litter fractions in an upland Amazonian Forest in Roraima, Brazil.c |
|
|||||||||||
|
|
|
|
|
|
|
|
||||||
|
Fraction |
Element |
|||||||||||
|
|
|
P |
|
K |
|
N |
|
Ca |
|
Mg |
C |
|
|
|
(kg ha-1
year-1) |
(%)* |
(kg ha-1
year-1) |
(%)* |
(kg ha-1
year-1) |
(%)* |
(kg ha-1
year-1) |
(%)* |
(kg ha-1
year-1) |
(%)* |
(kg ha-1
year-1) |
(%)* |
|
Leaves |
3.62 |
(0.065) |
26.76 |
(0.481) |
88.67 |
(1.592) |
35.61 |
(0.639) |
10.00 |
(0.180) |
2806.10 |
(50.38) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Wood |
0.62 |
(0.025) |
6.59 |
(0.271) |
25.55 |
(1.051) |
15.59 |
(0.642) |
2.20 |
(0.090) |
1090.20 |
(44.86) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Reproductive structures |
0.31 |
(0.110) |
2.28 |
(0.809) |
4.91 |
(1.743) |
0.68 |
(0.241) |
0.42 |
(0.149) |
135.40 |
(48.06) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Unclassified material |
0.95 |
(0.107) |
4.52 |
(0.508) |
18.97 |
(2.134) |
4.18 |
(0.470) |
1.44 |
(0.162) |
421.70 |
(47.43) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
TOTAL |
5.49 |
(0.060) |
40.15 |
(0.437) |
138.10 |
(1.506) |
56.07 |
(0.611) |
14.06 |
(0.153) |
4453.40 |
(45.56) |
|
(*) ‑ Values in
parentheses are concentrations (%) |
|
|
|
|
|
|
||||||
|
TABLE 3. Litter production and nutrient fluxes in upland
tropical forests of Brazilian Amazonia and neighboring parts of South America |
|
|
|||||||||
|
Location |
|
Production |
Nutrient flux (kg ha-1
year-1) |
Source |
|||||||
|
|
|
Total |
Leaves |
|
N |
P |
K |
Ca |
Mg |
|
|
|
|
|
(t ha-1 year-1) |
(t ha-1 year-1) |
(%) |
(t ha-1 year-1) |
(t ha-1 year-1) |
(t ha-1 year-1) |
(t ha-1 year-1) |
(t ha-1 year-1) |
|
|
|
BRAZILIAN AMAZONIA: |
|
|
|
|
|
|
|
|
|||
|
Manaus (Amazonas) |
7.40 |
5.60 |
(75.7) |
105.6 |
2.2 |
12.7 |
18.4 |
12.6 |
Klinge & Rodrigues
1968a,b |
||
|
Belém (Pará) |
9.90 |
8.00 |
(80.8) |
156.5 |
4.1 |
17.0 |
32.8 |
26.8 |
Klinge
1977b; Franken et al. 1979 |
||
|
Manaus (Amazonas) |
7.90 |
6.40 |
(81.0) |
‑ |
‑ |
‑ |
‑ |
‑ |
|
||
|
Tucuruí (Pará) |
6.65 |
4.76 |
(71.6) |
126.3 |
3.4 |
26.3 |
49.5 |
14.4 |
Silva 1984 |
||
|
Manaus (Amazonas)(a) |
8.25 |
5.42 |
(65.7) |
151.0 |
3.1 |
15.0 |
36.7 |
13.8 |
Luizão 1989 |
||
|
Manaus (Amazonas)(b) |
7.44 |
4.68 |
(63.0) |
109.0 |
3.7 |
22.2 |
58.2 |
14.0 |
Luizão 1989 |
|
|
|
Capitão Poco (Pará) |
8.04 |
|
‑ |
115.0 |
3.6 |
28.5 |
114.2 |
15.9 |
Dantas & Phillipson 1989 |
||
|
Maracá (Roraima) |
9.28 |
6.30 |
(67.9) |
117.9 |
6.7 |
48.5 |
63.7 |
23.8 |
Scott et al. 1992 |
||
|
Apiaú (Roraima) |
9.17 |
5.57 |
(60.7) |
138.1 |
5.5 |
40.2 |
56.1 |
14.1 |
This study |
||
|
NEIGHBORING PARTS OF SOUTH
AMERICA: |
|||||||||||
|
Colombia(c) |
12.02 |
6.51 |
(54.0) |
141.0 |
4.2 |
16.6 |
89.9 |
20.5 |
Folster
& de las Salas 1976; Folster et
al. 1976 |
||
|
Colombia(d) |
8.73 |
6.64 |
(73.0) |
|
|
|
|
|
Folster
& de las Salas 1976; Folster et
al. 1976 |
||
|
French Guiana |
8.64 |
5.78 |
(66.9) |
‑ |
‑ |
‑ |
‑ |
‑ |
Puig 1979 |
||
|
Venezuela(d) |
6.97 |
3.34 |
(48.0) |
69.0 |
4.0 |
33.0 |
43.0 |
14.0 |
Fassbender & Grimm 1981 |
||
|
Suriname |
|
9.30 |
|
‑ |
158.0 |
6.0 |
16.0 |
33.0 |
16.0 |
Ohler 1980; Poels 1987 |
|
|
Venezuela |
|
10.25 |
7.56 |
(73.8) |
167.1 |
3.1 |
24.6 |
17.4 |
7.2 |
Cuevas & Medina 1986 |
|
|
Colombia(e) |
7.03 |
4.61 |
(65.6) |
81.9 |
6.1 |
59.3 |
‑ |
‑ |
Veneklass 1991 |
||
|
Colombia(f) |
|
4.31 |
2.82 |
(65.4) |
34.2 |
1.9 |
13.9 |
‑ |
‑ |
Veneklass 1991 |
|
|
(a) Plateau forest; (b)
valley forest; (c) Includes 2.06 t ha-1 of palm fronds; (d)
Montane forest; (e) Andean forest 2550 m in altitude; (f) Andean forest 3370
m in altitude. |
|||||||||||
Fig. 1
Fig. 2